
Retina Regeneration

RPE Regeneration
RGC Neuroprotection
Retina Regeneration
Reza Raeisossadati - Postdoc
Vindya Senanayake - Ph.D. student
Adesola Johnson - Ph.D. student
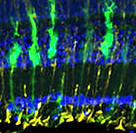
In fish and amphibians the retina is able to fully regenerate from a variety of insults but this ability is progressively lost in higher vertebrates, with modest regeneration in birds, and essentially no regeneration in mammals. Müller glia (MG) are responsible for retinal regeneration and due to the remarkable regenerative capacity of zebrafish, many studies have used this model to uncover the molecular underpinnings of retinal regeneration. In zebrafish, retinal damage first stimulates MG to adopt a gliotic state, but this is transient and they subsequently reprogram into a stem cell-like state, re-enter the cell cycle and generate multipotent neurogenic progenitors that then give rise to new neurons. In mammals, including humans, the initial MG gliotic response to damage is similar to that in zebrafish, but MG fail to reprogram or generate new neurons. Instead, gliotic responses persist and MG proliferate and eventually form a glial scar. Recent studies have demonstrated that ectopic expression of a key pro-regenerative gene identified in zebrafish, ascl1, in MG of the adult mouse retina, along with inhibition of histone deactylase (Hdac) activity, stimulates a regenerative response from normally non-regenerative MG. Regeneration correlated with increased chromatin accessibility at critical pro-regenerative loci and when combined with other published studies, support a model in which the epigenetic landscape regulating the expression of pro-regenerative genes during the injury and regenerative responses is a key regulator of the ability of MG to reprogram and stimulate regeneration. Our knowledge of the epigenetic regulation of retinal regeneration is limited and this has become a recent focus of experiments in the lab. Indeed, given that regenerative responses can be stimulated from normally non-regenerative MG in the mammalian eye, understanding the epigenetic regulation of retinal regeneration could be transformational in supporting the development of new therapeutics aimed at restoring neurons lost to retinal degenerative diseases and retinal injuries.

Cartoon of MG activity after a retinal injury (here, ablation of UV cones).
Current projects focus on understanding the role of epigenetic regulators like the Brd family of proteins (Lee et al., 2023), whether tet-family DNA hydroxymethyltransferases participate in retinal regeneration and understanding the cell biological underpinnings of Muller glia activation.

Brd4-dependent recognition of acetylated histones is required for regeneration of UV cones (see Lee et al., 2023) for details
RPE Regeneration
Lyndsay Leach - Research Assistant Professor
Donghwa Suh - Ph.D. Student
Suzie Green - Ph.D. Student
Chelsea Smith - Ph.D. Student
Mo Mojoudi - Ph.D. Student
Diseases resulting in degeneration of the retinal pigment epithelium (RPE) are among the leading causes of blindness worldwide, and no therapies exist that can replace RPE to restore vision. The RPE is a polarized monolayer of cells that separates the retina from the choroid and performs many critical functions for vision (reviewed in Strauss, 2005). Microvilli extend from the apical RPE surface and interdigitate with photoreceptor outer segments (POS), enabling the RPE to support photoreceptor health. The basal surface of the RPE abuts and helps to form Bruch’s membrane (BM), which, along with tight junctions between RPE cells, creates the blood-retina barrier and facilitates nutrient and ion transport between the retina and choriocapillaris. Additionally, RPE pigment prevents light scatter by absorbing stray photons.
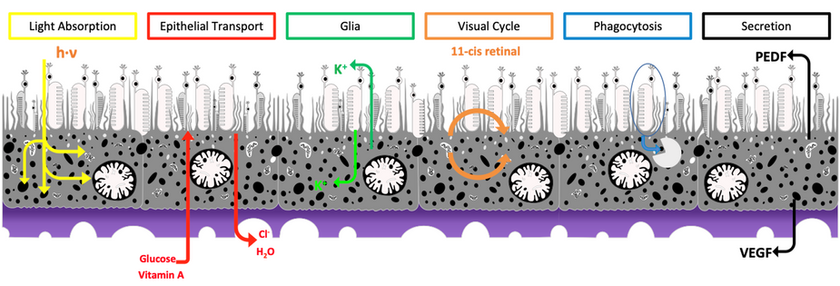
Cartoon of various RPE functions (adapted from Strauss, 2005).
Age-related macular degeneration (AMD) is a major RPE disease, and is the third leading cause of blindness in the world. AMD is commonly divided into two types: atrophic (dry) and exudative (wet). In the early stages of atrophic AMD, RPE cells in the parafovea become dysfunctional and progressively degenerate, and this is thought to result in death of parafoveal rod photoreceptors. Progressively, RPE dysfunction and degeneration spreads to the fovea, resulting in loss of cone photoreceptors and, ultimately, loss of high-acuity vision. Exudative AMD occurs in a subset of atrophic AMD cases when choroidal vasculature invades the retina.
Transplantation of stem cell-derived RPE has emerged as a possibility for treating AMD, with clinical trials are underway. An intriguing alternative approach to treat AMD and other RPE diseases is to develop therapies focused on stimulating endogenous RPE regeneration. For this to be possible, we must first gain a deeper understanding of the mechanisms underlying RPE regeneration and this has been a focus of our work over the last decade (reviewed in George et al., 2021 ). In mammals, RPE regeneration is extremely limited; small lesions can be repaired by the expansion of adjacent RPE cells, but remaining cells are unable to functionally replace lost RPE when the lesions are large. In some injury paradigms, RPE cells proliferate but do not regenerate a morphologically normal monolayer. A subpopulation of quiescent human RPE stem cells can be induced to proliferate in vitro and differentiate into RPE or mesenchymal cell types, but it is still unclear whether these exist in vivo.
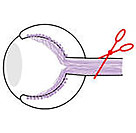
Despite these studies, little is known about the process by which RPE cells respond to injury to elicit a regenerative, rather than pathological, response. There are also few models in which to study intrinsic RPE regeneration. This knowledge gap is a major barrier to developing effective strategies to restore RPE lost to disease or injury. To overcome this barrier, we developed a transgenic zebrafish model to study RPE injury and regeneration (Hanovice et al., 2019).
We then leveraged this model to begin to identify the molecular and cellular mechanisms facilitating RPE regeneration.
Through our studies, we:
i) demonstrated that innate immune system activity is a critical regulator of RPE regeneration (Leach et al., 2021; 2025);
ii) showed that mTOR signaling modulates innate immune system activity during RPE regeneration (Lu et al., 2022) and
iii) performed a screen to identify novel regulators of RPE regeneration (Lu et al., 2023 ).

RPE ablation paradigm used in our studies.
Developed in collaboration with Ross Collery; see Hanovice et al., 2019 for details).
Our ongoing work is utilizing lineage tracing strategies to identify the source of regenerated RPE and we have performed single cell RNA sequencing to identify subpopulations of RPE cells pre/post injury. We’re also performing detailed morphometric analyses of normal RPE and after regeneration to better understand the regenerative process. Finally, we’re analyzing a number of genes and pathways identified through our screens and sequencing analyses to determine how they function during the regenerative process.
RGC Neuroprotection
Mia Pacheco - Postdoc
Retinal ganglion cells (RGCs) are the sole projection neurons connecting the retina to the brain and therefore play a critical role in vision. Death of RGCs during glaucoma, optic neuropathies and after ocular trauma results in irreversible loss of vision as RGCs do not regenerate in the human eye. Moreover, there are no FDA approved therapies that prevent RGC death and/or promote RGC survival in the diseased or injured eye. There is a critical need to better understand the molecular underpinnings of neuroprotection to develop effective therapeutic approaches to preserve damaged RGCs. Unlike in mammals, RGCs in zebrafish are resilient to optic nerve injury, even after complete transection of the optic nerve. Our work has identified a key role for Jak/Stat modulation of innate immune responses in this process (Chen et al., 2022).

More recently, we leveraged this unique model and utilized single-cell RNA sequencing to characterize RGC responses to injury and identify putative neuroprotective and regenerative pathways (Ali et al., 2025 ). RGCs are heterogeneous and studies in mice have shown that there is differential resiliency across RGC subtypes. Our results demonstrated that all RGC subtypes are resilient to injury in zebrafish. Quantifying changes in gene expression revealed the upregulation of progenitor and regenerative markers in all RGC subtypes after injury as well as distinct early and late phases to the injury response. This shift in gene expression causes injury-responsive RGCs to resemble RGC subtype 3, a low frequency population of endogenous immature RGCs that are normally maintained in the wild-type, uninjured adult retina. A similar but restricted transcriptomic injury response in RGCs of the uninjured contralateral eye was also detected, highlighting a systemic RGC response to unilateral optic nerve injury. Taken together, these results demonstrate that zebrafish RGCs dedifferentiate in response to injury, and this may be a novel mechanism mediating their unique cell survival and regenerative capabilities. Current work is focused on understanding this response at the molecular and genomic levels.

Single cell transcriptional analysis identifies molecular RGC subtype identities (see Ali et al., 2025 for details).